Social jetlag carries a measurable overnight autonomic cost across 187,796 ring wearers, sharply age-graded and largest in young women
Ultrahuman looked at data from nearly 188,000 users and found that people tend to go to bed about 24 to 34 minutes later on weekends compared to weekdays. And alongside that shift, overnight heart rate also went up.
The construct of social jetlag, the offset between sleep timing on free days and on work days, was named twenty years ago to describe a circadian misalignment that arises in healthy adults whose social schedules disagree with their endogenous chronotype (Wittmann et al., 2006). The construct is conventionally measured by Munich Chronotype Questionnaire (MCTQ) recall: the difference between the midpoint of sleep on free days and on workdays, in hours. A 65,000-respondent MCTQ analysis published six years later established that social jetlag is sharply age-graded, peaking in late adolescence and early twenties and declining steadily across the adult lifespan (Roenneberg et al., 2012).
What was not previously measurable at scale was the autonomic cost of that misalignment in vivo, on consumer-wearable signal, in a cohort large enough to resolve effect sizes within sex and age strata. Across 187,796 long-term Ultrahuman ring wearers in Western timezones over four months, social jetlag tracks a small but uniform rise in overnight resting heart rate and a fall in overnight HRV, with both effects more than twice as large in 18-to-29-year-old women as in 50-to-75-year-old women.
A mechanistic context for that pattern has been gathering. A 360-adolescent Penn State Child Cohort study using 24-hour Holter-derived HRV reported in 2025 that later free-day sleep midpoints blunted every one of five overnight HRV measures tested (Morales-Ghinaglia et al., 2025). A 2023 review traced the upstream physiology to enhanced sympathetic drive and disrupted parasympathetic withdrawal during sleep when the circadian phase is misaligned with the timing of behavioural rest (Shafer et al., 2023). A 2026 analysis of 20,968 wearable users built on the same body of work, reporting a within-person dose-response of nocturnal RHR rise and HRV fall per drink above each user's personal mean, with effects amplified in women and in adults under 35 (Grosicki et al., 2026). What that paper could not directly resolve was how much of the weekend autonomic shift is driven by alcohol exposure specifically versus the broader timing misalignment that often co-travels with weekend behaviour. The analysis here addresses the timing axis directly: it measures the social-jetlag exposure as a behavioural variable derived from ring-recorded sleep midpoints, with no claim about which downstream behaviours (alcohol, late eating, deferred exercise, reduced morning activity) drive the autonomic delta.
The cohort definition was deliberately tight on three axes. Eligible members were ring wearers with at least 20 valid work-night sleep records and at least 8 valid free-night records in the 120-day window from 23 December 2025 to 22 April 2026, where validity required the index-valid flag, age between 18 and 75, and gender recorded as 0 (male) or 1 (female); internal users were excluded at the SQL level on every query. Members in Asia/* timezones were excluded by pre-registered design rather than post-hoc, on the grounds that the Friday-Saturday weekend convention on which the MCTQ free-day-versus-work-day mapping rests does not generalise to the dominant Ultrahuman user base in India and East Asia. The resulting cohort split into 8 age-by-gender cells, the smallest of which (40-to-49-year-old men) carried 10,695 members; the largest (18-to-29-year-old women) carried 74,577. Per-user social jetlag was operationalised as the median sleep-midpoint on free nights (Friday and Saturday evenings local) minus the median midpoint on work nights, with the midpoint expressed as hours past 18:00 local to avoid midnight-wrap artefacts. Outcome deltas were computed within-person: the difference between mean overnight resting heart rate (and mean overnight HRV) on free-night mornings versus work-night mornings.
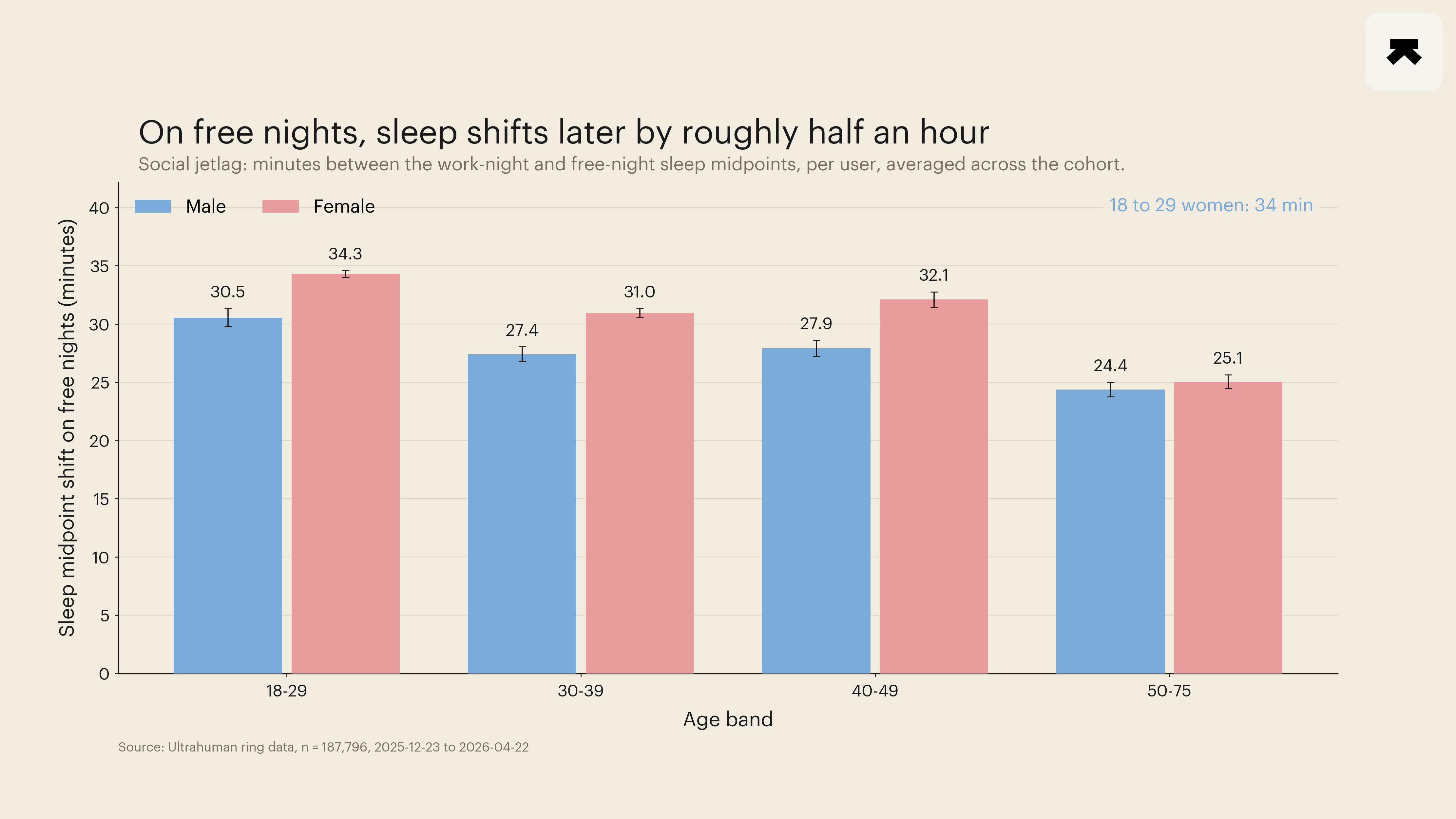
Three patterns surface. First, the exposure itself reproduces the Roenneberg age gradient.
Mean per-user social jetlag ran 34.3 minutes in 18-to-29-year-old women, fell to 25.1 minutes in 50-to-75-year-old women, and tracked a similar but flatter trajectory in men (30.5 minutes at 18-29; 24.4 at 50-75). Within every age band, women carried 3 to 5 minutes more social jetlag than men.
The MCTQ-recall gradient and the ring-PPG-timing gradient agree on direction and on rank-order across cells.
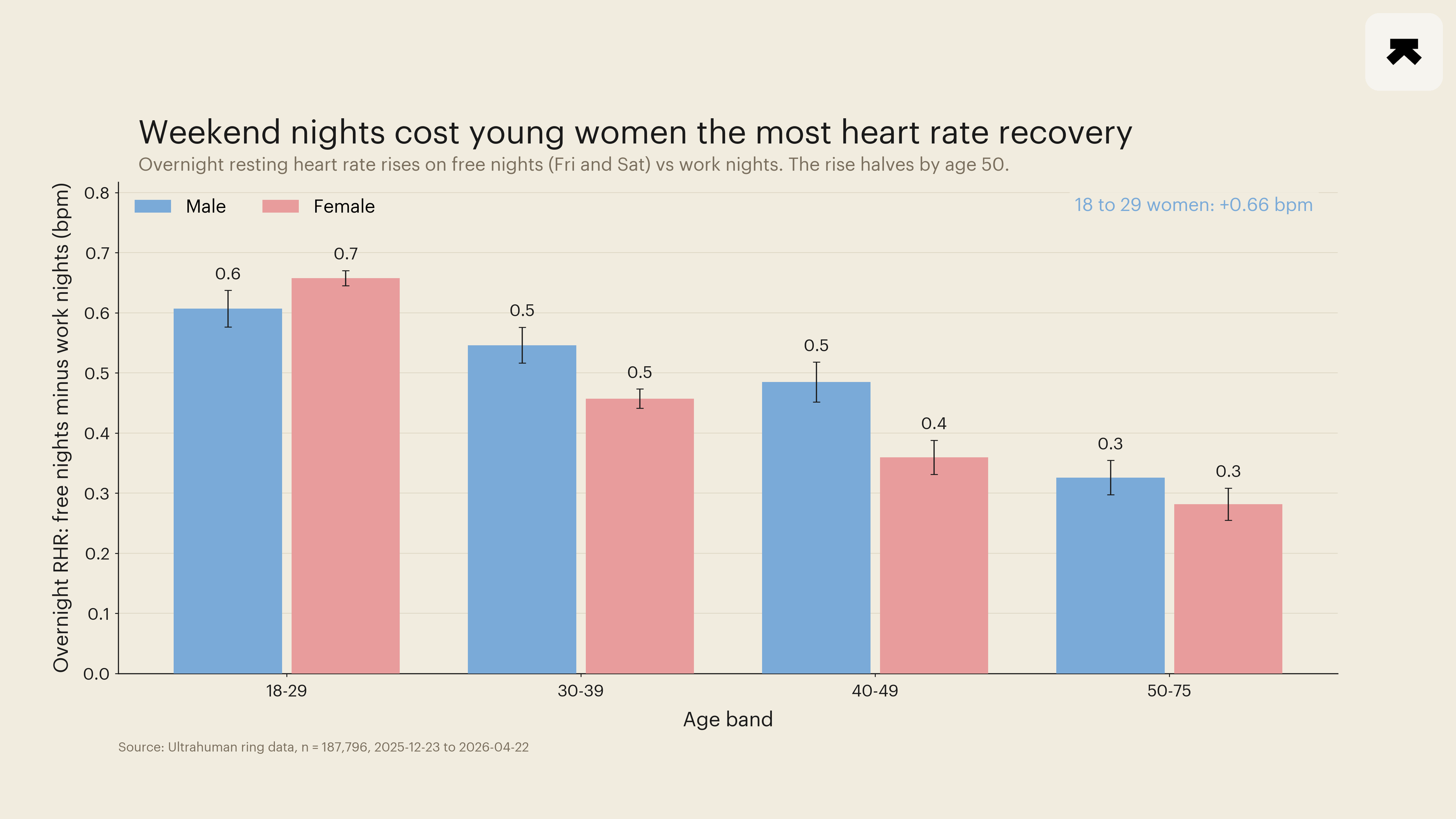
Second, the autonomic cost mirrors the exposure. Free-night minus work-night overnight RHR was positive in every one of the 8 cells, with every 95 percent confidence interval clearing zero. The cell extremes were +0.66 bpm in 18-to-29-year-old women (CI +0.64 to +0.67) and +0.28 bpm in 50-to-75-year-old women (CI +0.26 to +0.31), a 2.4-fold age gradient with the youngest and oldest female CIs not touching. Free-night minus work-night overnight HRV was negative in every cell on the same gradient, with the youngest women dropping 0.87 ms and the oldest women 0.32 ms. The two autonomic markers move in their expected opposite directions (rise in mean nocturnal heart rate; fall in beat-to-beat variability) on the same age structure.
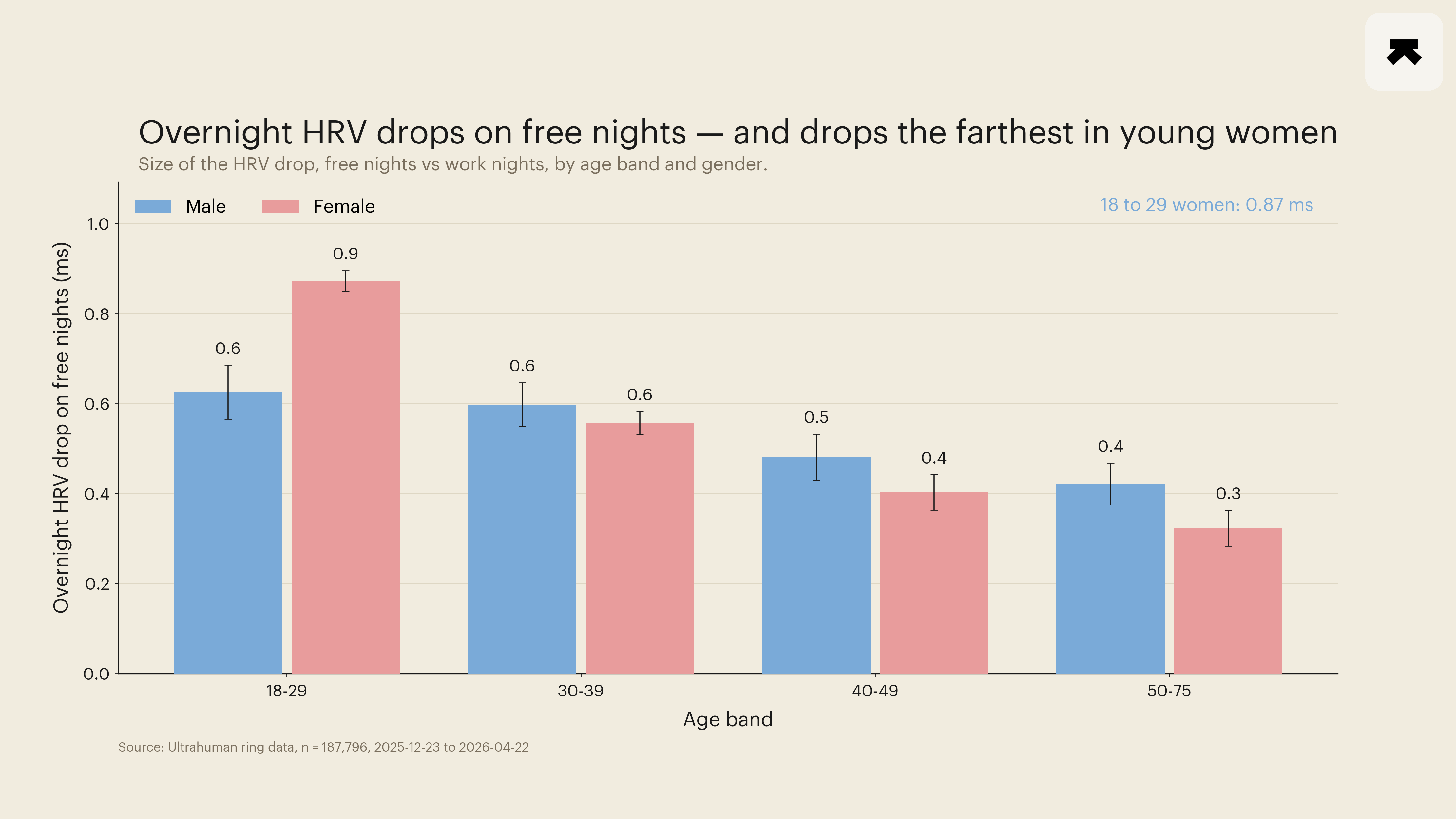
Third, the gender amplification predicted by the alcohol-exposure literature appears here as well, but only in the youngest band. The female-versus-male difference within the 18-to-29 cell is +0.05 bpm on RHR and -0.24 ms on HRV, with the female direction always larger in magnitude. The female-versus-male gap closes by the 30-39 band on RHR (males show +0.55, females +0.46) and stays modest through 50-75 (males +0.33, females +0.28). This is the place where the wearable-cohort observation departs informatively from the alcohol-paper headline: the sex amplification appears robustly in the youngest stratum and not as a uniform across-age effect, which is consistent with younger adults being the population whose social-jetlag exposure is highest and whose drinking and late-eating co-travellers are most concentrated.
Two pre-registered stability checks were run. The 8-cell pattern was re-estimated on two non-overlapping 60-day sub-windows (winter 23 December to 21 February; spring 22 February to 22 April) with looser inclusion (at least 10 work-nights and 4 free-nights per sub-window). The sign and rank-order of every cell in every primary comparison reproduced. Magnitudes were larger in the spring window than the winter window: 18-to-29-year-old female RHR delta ran +0.81 bpm in spring versus +0.53 bpm in winter, and the corresponding HRV deltas were -1.12 ms and -0.65 ms. Whether that magnitude difference reflects daylight-driven seasonal physiology, behavioural changes (longer evenings, more outdoor late-week activity in spring) or some interaction is not resolvable from this analysis. The pooled-window estimates are reported as the headline; the sub-window estimates are reported as a transparency check. The age gradient and sex amplification reproduce in both.
Several caveats sit on this read and are worth naming directly. The free-day construct here is calendar-based (Friday and Saturday evenings) rather than alarm-use-based; the full MCTQ chronotype-corrected social-jetlag metric (MSFsc) cannot be computed from ring telemetry without per-user alarm tagging. The exclusion of Asia/* timezones means the result generalises to Western-timezone Ultrahuman members and not to the dominant Indian user base. The autonomic deltas are not adjusted for behavioural co-travellers (alcohol, late dinner, screen exposure, reduced morning activity); they describe the aggregate weekend autonomic footprint visible to the ring, not the contribution of any single behaviour to that footprint. Within-person menstrual-cycle variation in HRV is not corrected; given the within-cell aggregation across tens of thousands of women-nights and the random alignment of weekend nights with cycle phase across the cohort, cycle effects should average out at this scale, but the absence of a per-user correction is a methodological limit. Finally, the design is observational and the language is associational: there is no causal claim that intervening on bedtime regularity would shift overnight cardiovascular markers in the magnitudes observed here.
What the data say is narrow and falsifiable. In a long-term Western-timezone ring cohort, social-jetlag exposure reproduces the Roenneberg age gradient at MCTQ-comparable magnitudes and tracks an autonomic cost on overnight resting heart rate and HRV that is strongest in the youngest age band, sharply attenuates by 50, is slightly amplified in young women relative to young men, and reproduces across two seasonally distinct sub-windows. That is consistent with the construct being measurable on consumer ring data at population scale, and with the literature's mechanistic frame of misalignment driving sympathetic activation rather than alcohol exposure alone driving the weekend physiology.
- Wittmann M, Dinich J, Merrow M, Roenneberg T. Social jetlag: misalignment of biological and social time. Chronobiology International, 2006. PMID: 16687322.
- Roenneberg T, Allebrandt KV, Merrow M, Vetter C. Social jetlag and obesity. Current Biology, 2012. PMID: 22578422.
- Morales-Ghinaglia N, et al. Social jetlag and 24-hour heart rate variability in adolescents: the Penn State Child Cohort. Sleep, 2025. PMID: 39826092.
- Shafer BM, Kogan SA, McHill AW. Pressure building against the clock: recent advances in understanding the consequences of circadian misalignment on cardiovascular physiology. Current Hypertension Reports, 2023. PMID: 37837518.
- Grosicki GJ, et al. Within-person changes in alcohol consumption are associated with diminished objective sleep quality, autonomic nervous function, and physical activity in 20,968 wearable device users. PLOS Digital Health, 2026. PMID: 41801993.